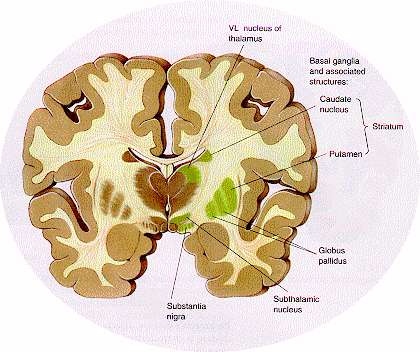
기저핵이란? 대뇌피질 밑의 백질 내에 있는 신경세포핵 덩어리로서 소뇌 피질 척수계 및 뇌간 운동핵들과 더불어 운동조정 기능에 참여한다.
@기저핵의 성분 (현재의 개념)
선조체(corpus striatum)
선조(striatum)
미상핵(caudate nucleus)
조가비핵(putamen, 피각)
배쪽선조(ventral striatum) - 중격의지 핵(nucleus accumbens septi)
창백(pallidum, 담창)
창백핵(globus pallidus, GP, 담창구)
내핵(내측분절, internal or medial segment, GPi)
외핵(외측분절, external or lateral segment, GPe)
배쪽창백(ventral pallidum) - 무명질(substantia innominata)의 일부
흑색질(substantia nigra, 흑질)
그물부(pars reticulata, SNr, 망상부)
치밀부(pars compacta, SNc, A9)
시상밑핵(subthalamic nucleus, 시상하핵)
@선조 (Striatum)
선조(striatum), 또는 신선조(neostriatum)는 미상핵(caudate nucleus)과 조가비핵(putamen)으로 구성되어 있으며, 앞쪽으로는 내섬유막 전각의 아래쪽에서 연속되 어 있고, 미상핵-렌즈핵 회색질 연결부(caudatolenticular gray bridge)에 의해서도 연결되어 있다. 이 두 핵의 형태는 거의 동일하다. 세포는 매우 많고 밀집되어 있으며, 옅은 회색으로 나타나는 창백 핵과는 달리 혈관이 많아 약간 붉은 빛을 띤 회색의 구조로 나타난다. 또한 두꺼운 유수섬유다발 이 가로지르는 창백핵과는 달리 가는 유수섬유와 무수섬유의 다발이 이 핵으로 들어온다.
1. 선조신경원(Striatal Neuron)
이 두 핵을 이루는 신경세포의 형태는 거의 동일하다. 대부분의 세포는 작은 다극신경원으로 구성되어 있으며, 일부 큰 다극신경원이 존재한다. 골지염색에서 나타나는 선조의 신경원은 크게 긴 투사성 축삭(long projecting axon)이 있는 신경원과 짧은 축삭이 있는 신경원으로 나눌 수 있다.
㈀축삭이 긴 투사성 신경원: 수상돌기(dendrite)에 가시(spine, 소극)가 있기 때문 에 소극세포(spiny cell)라고 불리우며 다시 제1형소극세포(type I spiny cell, spiny I cell, S I)와 제2 형소극세포(type II spiny cell, spiny II cell, S II)의 두 종류로 나누어진다.
①제1형소극세포(S I): 선조 신경원의 대부분(96%)을 차지하고 있으며 세포체는 중간 정도 크기이고 축삭은 창백(GP)과 흑색질(substantia nigra)로 투사된다. 신경전달물 질로 억제성인 감마아미노부티르산(GABA)를 함유하고 있으며, 엔케팔린(enkephalin)과 섭스탄스 피(substance P)를 공유하는 경우도 있다고 알려져 있다. 수상돌기의 근위부(세포체에서 20 ‡ 6;m 정도)에는 소극이 없으나 원위부에는 소극이 매우 촘촘하게 밀집되어 있다. 수상돌기는 여러 방향으로 뻗어 있으며 대체로 세포체를 중심으로 직경 200 ?m 정도의 공간을 차지하고 있 다.
피질선조섬유(corticostriate fiber)와 시상선조섬유(thalamostriate fiber)는 주로 이 세포의 수상돌기소극(dendritic spine)과 시냅스를 이룬다. 이 두 가지 구심섬유는 모두 흥분성이며, 피질선조섬유의 신경전달물질은 글루탐산(glutamic acid)라고 알려져 있고 시상선조섬유의 신경전 달물질도 글루탐산으로 추측되고 있다. 소극이 없는 축삭근위부와 세포체에는 주로 선조의 중간 신경원(interneuron)에서 아세틸콜린(acetylcholine), 섭스탄스 피(substance P), 감마아미노부티르산(G ABA) 등을 함유한 축삭종말이 연접한다. 소극이 있는 수상돌기 원위부에서 소극 사이에는 도파 민(dopamine)을 신경전달물질로 하는 흑색질 치밀부(SNr)에서 들어오는 흑색선조섬유가 시냅스를 이룬다.
원심섬유는 주로 창백(GP)과 흑색질 그물부(SNr)로 투사되며, 일부 원심섬유는 흑색질 치밀부(SNc)로도 투사된다. 제1형소극세포 축삭의 곁가지는 다른 제1형소극세포에도 연접 된다고 알려져 있다(그림 9-5). 이러한 곁가지는 생리 학적으로 주위의 세포의 전달과정을 억제하는 외측억제(lateral inhibition)의 기능을 한다고 추측된 다. 창백외핵(GPe)으로 투사되는 신경원은 주로 감마아미노부티르산(GABA)과 엔케팔린(enkephali n)을 신경전달물질로 공유하고 있으며, 창백내핵(GPi)과 흑색질 그물부(SNr)로 투사되는 신경원은 주로 감마아미노부티르산(GABA)과 섭스탄스 피(substance P)를 공유하고 있는 것으로 보고되어 있 다. 흑색질 치밀부(SNr)로 투사되는 신경원은 선조소체(striosome)의 신경원이다(선조구획 참조).
②제2형소극세포(S II): 선조 신경원의 약 1%를 차지하고 있으며 세포 체는 비교적 크고 방추형이다. 수상돌기도 더 멀리까지 뻗어 있어 직경 600 ?m 정도의 공 간을 차지하고 있다. 수상돌기의 어느 부분에나 수상돌기가시가 있으나 제1형세포에 비해서는 드 물게 나타난다. 축삭은 창백(GP)과 흑색질 그물부(SNr)로 투사되며, 흥분성인 섭스탄스 피(substanc e P)를 신경전달물질로 함유하고 있다고 알려져 있다
㈁축삭이 짧은 신경원
수상돌기에 가시가 없기 때문에 무극세포(aspiny cell)라 고 하며 다시 제1형무극세포(type I aspiny cell, aspiny I cell, A I), 제2형무극세포(type II aspiny cel l, aspiny II cell, A II), 제3형무극세포(type III aspiny cell, aspiny III cell, A III)의 세 종류로 세분 된다.
①제1형무극세포(A I) :역시 선조 신경원의 1%를 차지하고 있으며 세포 체는 비교적 작다. 신경전달물질로는 GABA와 NPY를 함유하고 있을 것이라고 추측되고 있다. 축 삭은 제2형무극세포에 억제성 연접을 형성한다고 알려져 있다.
②제2형무극세포(A II):세포체가 매우 큰 세포로 선조 신경원의 1% 정도를 차지하고 있다. 신경전달물질로 흥분성인 아세틸콜린(acetylcholine)을 함유하고 있다. 흑색 질 치밀부(SNc)에서 기원된 도파민성 섬유는 이 세포의 활성을 강력하게 억제한다고 알려져 있 다. 이 세포의 축삭은 제2형소극세포에 흥분성 연접을 형성한다
③제3형무극세포(A III):선조 신경원의 약 1%를 차지하고 있으며 세 포체는 작고 어떠한 종류의 신경전달물질을 함유하고 있는지는 분명하지 않다. 제2형소극세포(S I I)에서 구심성 섬유를 받으며 원심성 섬유는 다시 제2형소극세포에 억제성 연접을 형성한다고 알 려져 있다
<그림에 관한 설명>
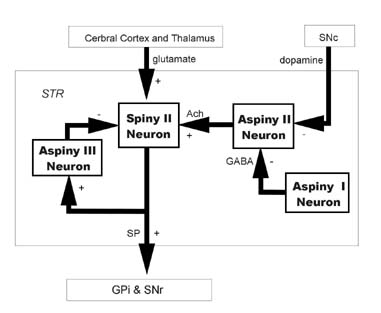
제2형소극세포(type II spiny cell, S II)의 연결회로. 제2형소극세포(S II)도 대뇌피질과 시상에서 흥분성 입력을 받지만, 제1형소극세포(S I)와는 달리 신경전달물질로 substance P를 함유하고 있고, 창백(GP)과 흑색질 그물부(SNr)로 흥분성 출력을 보낸다. 이 세포(S II)는 아세틸콜린(Ach)을 신경전달물질로 함유한 제2형무극세포(A II)에서 흥분성 입력을 받으며, 제2형무극세포(type II aspiny cell, A II)는 도파민성인 흑색질 치밀부(SNc)에서 강한 억제성 입력을 받는다. 제3형무극세포(type III aspiny cell, A III)는 창백(GP)과 흑색질그물부(SNr)로 가는 제2형소극세포(S II)의 곁가지에서 흥분성 입력을 받으며, 다시 제2형소극세포(S II)로 억제성 출력을 보낸다. 제1형무극세포(type I aspiny cell, A I)는 신경전달물질로 억제성인 감마아미노부티르산(GABA)를 함유하고 있어, 제2형무극세포(A II)로 억제성 출력을 보낸다.
2. 선조의 연결 (Striatal Connections)
기저핵 중에서 선조(striatum)는 여러 부위에서 구심성 섬유를 받아 이를 출력부(창백 GP 과 흑색질 그물부 SNr)로 보내주는 입력부(input portion)라고 할 수 있다.
㈀구심성 연결 (afferent connections) :
선조는 대뇌피질(cerebral cortex), 시상(thalamus)의 수질판내핵군(intralaminar nuclear group), 흑색질 치밀부(SNc), 편도체(amygdaloid body), 뇌간의 솔기핵(raphe nuclei) 등 여러 구 조에서 구심섬유를 받는다.. 이러한 구심섬유 들은 도파민, 세로토닌, 글루탐산 등 여러 가지 신경전달물질을 함유하고 있는 것이 특징이다.
ⓛ피질선조로(corticostriate fiber): 대뇌피질에서온다.
거의 모든 신피질(neocortex)에서 기원되며 부위별 대응연결을 하고 있다. 특히 조가비핵(putamen)은 전두엽(frontal lobe)의 중심전이랑(precentral gyrus)에서 대부분의 구심성 섬유를 받으며, 미상핵(caudate nucleus)은 전전두엽피질(prefrontal cortex)에서 주로 구심섬유를 받는다. 피질선조로는 신경전달물질로 흥분성인 글루탐산(glutamic acid)을 함유하고 있다. 피질선조로의 신경원은 주로 대뇌피질의 내피라미드 층(internal pyramidal layer)에서 기원된다. 해마체(hippocampal formation)와 편도체(amygdaloid body)등 변연계(limbic system)에 속하는 구조에서는 주로 배쪽선조(ventral striatum)인 중격의지핵(nucleus accumbens septi)으로 신경섬유를 보내며, 주위의 미상핵 머리와 조가비핵으로도 투사섬유를 보낸다.
②시상선조로(thalamostriate fiber):시상의 수질판내핵군에서 나온다
주로 중심정중핵-다발옆핵 복합체(CM-PF complex)에서 기원되며, 역시 부위별로 대응연결되어 있다. 시상의 이 부위는 기저핵의 창백내핵(GPi)으로부터 구심섬유를 받는다. 흥분성으로 작용하며, 신경전달물질은 글루탐산으로 추측되나 아직 확실하게 알려지지 않았다.
③흑색선조로(nigrostriate fiber): 흑색질의 치밀부(SNc)에서 기원된다.
대부분 신경전달물질로 도파민(dopamine)을 함유하고 있다. 이 섬유는 선조의 제2형무극세 포(A II)를 억제하는 작용을 한다고 알려져 있다. 모든 흑색선조섬유가 도파민을 함유하고 있지는 않으며, 약 20% 정도는 도파민을 함유하고 있지 않다고 알려져 있다.
④편도선조로(amygdalostriate fiber): 편도체의 기저외측핵(basolateral nucleus)에서 기원된다.주로 배쪽선조(ventral striatum)인 중격의지핵(nucleus accumbens septi)으로 신경 섬유를 보내며, 미상핵 머리의 배쪽내측부와 조가비핵의 배쪽내측부로도 투사섬유를 보낸다

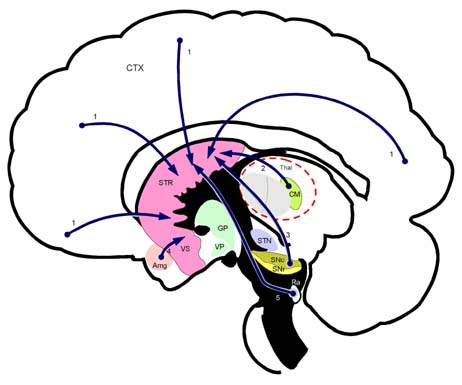
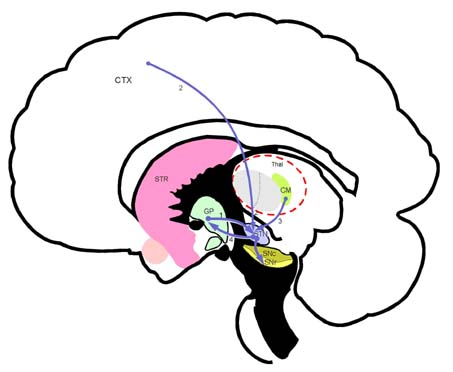
Amg 편도체(amygdaloid body) CM 중심정중핵(centre median nucleus)
Ra 솔기핵(raphe nuclei, 봉선핵) GP 창백핵(globus pallidus, 담창구)
CTX 대뇌피질(cerebral cortex) SNr 흑색질 그물부
SNc 흑색질 치밀부 STN 시상밑핵(subthalamic nucleus)
Thal 시상(thalamus) VS 배쪽시상(ventral striatum)
VP 배쪽창백(ventral pallidum) 1. 피질선조섬유(corticostriatal fiber)
2. 시상선조섬유(thalamostriatal fiber) 3. 흑색선조섬유(nigrostriatal fiber)
4. 편도선조섬유(amygdalostriatal fiber) 5. 솔기선조섬유(raphe striatal fiber)
선조(striatum, STR; 미상핵 caudate nucleus 과 조가비핵 putamen)의 구심성 연결(afferent connections). 선조(STR)는 기저핵(basal ganglia) 전체의 입력부(input portion)로 대뇌피질(CTX), 시상(Thal), 흑색질 치밀부(SNr), 편도핵복합체(Amyg), 솔기핵(Ra) 등에서 오는 구심섬유를 받는다.

㈁ 원심성 연결 (efferent connections)
선조에서는 기저핵의 출력부인 창백핵(GP)과 흑색질 그물부(SNr)로 원심성 섬유를 보낸다
선조에서 창백핵으로 투사되는 선조창백섬유(striopallidal fiber)는 질서정연하게 부챗살 모양으로 창백핵의 내핵(GPi)과 외핵(GPe)에 모두 종지하며, 이 섬유다발을 윌슨의 필모섬유다발(pencil bundle of Wilson)이라고도 부른다. 일부 선조 신경원은 흑색질 그물부(SNr)로 이어지는 선조흑색질섬유(strionigral fiber)를 형성한다. 원심섬유의 신경전달물질은 대부분 억제성인 감마아미노부티르산(GABA)이며, 엔케팔린(enkephalin)과 섭스탄스 피(substance P)를 공유하고 있는 신경원이 많다. 창백외핵(GPe)으로 투사되는 신경원은 주로 감마아미노부티르산(GABA)과 엔케팔린(enkephalin)을 신경전달물질로 공유하고 있으며, D2 도파민수용체를 함유한다. 창백내핵(GPi)과 흑색질 그물부(SNr)로 투사되는 신경원은 주로 감마아미노부티르산(GABA)과 섭스탄스 피(substance P) 및 다이놀핀(dynorphin)을 공유하고 있으며, D2 도파민수용체를 함유하고 있다고 보고되어 있다. 헌팅튼병(Huntington's disease)에서 처음으로 파괴되는 구조는 창백외핵(GPe)으로 투사되는 선조의 신경원이라고 알려져 있다. 일부 선조신경원은 기저핵의 출력부가 아닌 흑색질 치밀부(SNc)로 직접 원심섬유를 보낸다. 이러한 선조신경원은 선조소체(striosome)에 있으며, 일부 변연피질(limbic cortex)에서 구심섬유를 받는다.

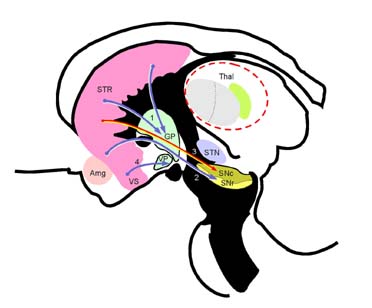
Amg 편도체(amygdaloid body) GP 창백핵(globus pallidus, 담창구)
SNc 흑색질 치밀부(substantia nigra, pars compacta)
SNr 흑색질 그물부(substantia nigra, pars reticulata)
STN 시상밑핵(subthalamic nucleus) Thal 시상(thalamus)
VS 배쪽시상(ventral striatum) VP 배쪽창백(ventral pallidum)
1. 선조창백섬유(striopallidal fiber) 2. 선조흑색질섬유(strionigral fiber)
3. 선조소체(striosome)에서 흑색질치밀부(SNc)로의 원심섬유
4. 배쪽선조배쪽창백섬유(ventrostriatoventropallidal fiber)
선조(striatum, STR; 미상핵 caudate nucleus 과 조가비핵 putamen)의 원심섬유(efferent fiber). 대부분의 선조원심섬유는 기저핵(basal ganglia)의 출력부인 창백(globus pallidus, GO)과 흑색질 그물부(pars reticulata of substantia nigra, SNr)로 가기 때문에 이 연결은 기저핵 입력부와 출력부의 연결이라고 할 수 있다. 선조소체(striosome)에서 출발하는 일부 선조원심섬유만이 출력부가 아닌 흑색질 치밀부(SNc)로 이어진다.

창백 (Pallidum)
1.창백의 구조와 신경세포
창백은 창백핵(globuspallidus)을 말하며, 조가비핵(putamen)의 내측에 위치한다. 외측으로는 외측수질판(external medullary lamina)에 의해 조가비핵과 경계지워지며, 내 측으로는 내섬유막(internal capsule)에 의해 간뇌(diencephalon)와 구분된다. 중간에는 내측수질판(int ernal medullary lamina)이 있어 창백내핵(내측분절, internal or medial segment, GPi)과 창백외핵(외측분절, external or lateral segment, GPe)으로 나누어진다. 하등 포유류에서 창백내 핵은 대뇌각내핵(entopeduncular nucleus)이라고 한다.
신경세포는 선조의 신경세포에 비해 크며, 산재되어 있다. 대부분 다극신경원 이며 수상돌기에는 수상돌기가시(dendritic spine)가 많지 않다. 약간 붉은 빛을 띠는 선조와 달리, 옅은 회색으로 나타난다. 두꺼운 유수섬유다발이 지나가므로 수초염색에서는 선조에 비해 짙게 염색되는 섬유다발이 관찰된다. 창백핵의 바로 아래에는 배쪽창백(ventral pallidum)을 포함 하는 무명질(substantia innominata)이 위치해 있다.
2.창백의 연결 (Pallidal Connections)
㈀구심성 연결 (afferent connections)
창백핵(GP)은 기저핵의 출력부로 선조(striatum)와 시상밑핵(subthalamic nucleus, STN)으로부터 구심섬유를 받는다
선조창백섬유(striopallidal fiber)는 부챗살 모양으로 창백핵의 내핵(GPi)과 외핵(GPe)에 모두 신경섬유를 보낸다. 창백외핵(GPe)으로 투사되는 섬유는 주로 감마아미노부티르산(GABA)과 엔케팔린(enkephalin)을 신경전달물질로 공유하고 있으며, 창백내핵(GPi)으로 투사 되는 신경원은 주로 감마아미노부티르산(GABA)과 섭스탄스 피(substance P) 및 다이놀핀(dynorphi n)을 공유하고 있다.
시상밑핵에서 기원되는 시상밑창백섬유다발(subthalamopallidal fiber bundle)은 선조창백섬유다발(striopallidal fiber bundle)에 비해 작다. 이들 섬유 역시 부위별 대응연결을 하고 있으며, 창백의 내핵(GPi)과 외핵(GPe) 모두에 종지한다. 반면 창백핵에서 시상밑핵으로 이어지는 창백시상밑다발은 창백외핵(GPe)에서만 기원한다.

시상밑핵(subthalamic nucleus)의 연결. 시상밑핵(STN)은 주로 창백외핵(GPe)에서 구심섬유(창백시상밑핵섬유 pallidosubtahalmic fiber)에서 구심섬유를 받으며, 대뇌피질, 시상의 중심정중핵-다발옆핵 복합체(CM- PF complex)에서도 구심섬유를 받는다. 시상밑핵(STN)의 원심섬유는 창백내핵(GPi), 창백외핵(GPe), 흑색질 그물부(SNr) 등 기저핵 출력부 전체에 투사섬유를 보낸다.
 .
.
㈁원심성 연결 (efferent connections)
창백에서는 시상, 시상밑핵(STN), 시상상부(epithalamus)의 외측고삐핵(lateral habenular nucleus), 뇌간의 대뇌각교뇌핵(pedunculopontine nucleus, PPN) 등 여러 부위로 원심성 섬유를 보낸다. 대부분의 섬유는 시상(thalamus)과 시상밑핵(STN)으로 투사된다

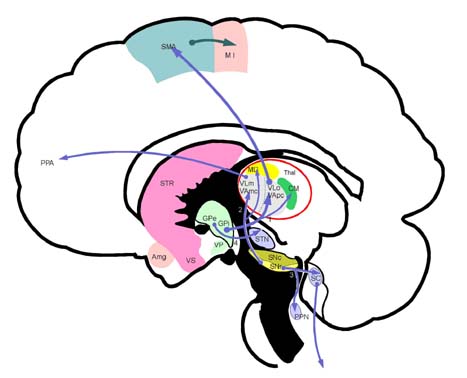
Amg 편도핵복합체 M I 일차운동영역 CM 중심정중핵
MD 등쪽내측핵 PPA 전전두엽 연합영역 Thal 시상
PPN 대뇌각교뇌핵 SC 상구 SMA 보완운동영역
STN 시상밑핵 GPi 창백내핵 SNc 흑색질 치밀부
STR 선조체 SNr 흑색질 그물부 GPe 창백외핵
VAmc 배쪽전핵거대세포부 VApc 배쪽전핵소세포부
VLm 배쪽외측핵내측부 VLo 배쪽외측핵 구부
VS 배쪽시상(ventral striatum) VP 배쪽창백(ventral pallidum)
1. 창백시상섬유(pallidothalamic fiber) 2. 흑색시상섬유(nigrothalamic fiber)
3. 흑색질피개섬유(nigrotectal fiber) 4. 창백시상밑섬유(pallidosubtahalmic fiber)
5. 덮개시상로(tectospinal tract)
기저핵(basal ganglia)의 원심신경로(efferent fibers). 기저핵의 출력부인 창백핵(globus pallidus)과 흑색질 그물부(SNr)에서는 시상과 시상하해 및 뇌간으로 원심섬유를 보낸다. 창백내핵(GPi)에서는 시상(VLo, VApc, CM), 으로 원심섬유를 보내며, 창백외핵(GPe)에서는 시상밑핵(STN)으로 원심섬유를 보낸다. 흑색질 그물부에서는 시상(VLm, VAmc)과 중뇌의 상구(superior colliculus), 대뇌각교뇌핵(pedunculopontine nucleus, PPN)으로 연결되어 있다. 창백내핵(GPi)과 흑색질 그물부(SNr)의 원심섬유는 시상의 각기 다른 핵으로 원심섬유를 보낸다.

@흑색질
흑색질은 교뇌중심회색질의 위쪽 끝부분에서 시상밑핵(subthalamic nucleus)이 나타나는 위치까지 거의 중뇌 전체에 걸쳐 있는 핵으로, 대뇌각기저부(crus cerebri)와 중뇌 피개(tegmentum) 사이에 위치해 있다.
흑색질은 피개쪽에 위치한 치밀부(pars compacta)와 대뇌각기저부쪽에 위치한 그물부(pars reticulata)의 두 부분으로 나누어진다.
1.흑색질그물부
그물부의 신경원들은 도파민을 함유하고 있지 않으며 세포들의 밀도가 낮다. 전체적으로 치밀부보다 약간 위쪽에 위치해 있다. 이 세포의 형태는 기저핵에 속하는 창백핵(globus pallidus)의 세포와 비슷하며 실제로 서로 이어져 있는 부분도 있다(사노구역 field of Sano). 신경전달물질로 감마-아미노부티르산(γ-aminobutyric acid, GABA)을 함유한 세포들이 많다.
2.흑색질 그물부(SNr)의 연결
㈀구심성 연결 (afferent connection)
흑색질 그물부(SNr)는 주로 선조(striatum)에서 구심섬유를 받으며, 시상밑핵(STN)과 뇌간의 솔기핵(raphe nuclei), 그리고 대뇌각교뇌핵(PPN)에서도 약간의 구심섬유를 받는다고 알려져 있다.선조에서 흑색질 그물부(SNr)로 이어지는 선조흑색질섬유(strionigral fiber)는 부위별 대응 연결이 뚜렷하여, 미상핵 머리의 신경원은 흑색질 그물부의 상부로 투사되고, 조가비핵의 등쪽부분은 흑색질의 외측, 조가비핵의 배쪽부분은 흑색질의 내측으로 투사된다. 선조흑색질섬유는 주로 감마아미노부티르산(GABA)과 섭스탄스 피(substance P) 및 다이놀핀(dynorphin)을 공유하고 있다고 보고되어 있다.
시상밑핵(STN)에서 들어오는 상당 수의 시상밑흑색질섬유(subthalamonigral fiber)는 시상밑창백섬유(subthalamopallidal fiber)와 함께 시상밑핵의 주원심경로 중의 하나이다.
㈁원심성 연결 (efferent connection)
흑색질 그물부(SNr)의 원심섬유는 시상의 외측배쪽핵-배쪽전핵(VL-VA)과 등쪽내측핵(MD), 상구(superior colliculus), 및 대뇌각교뇌핵(PPN)으로 연결된다.
①흑색질시상섬유(nigrothalamic fiber)
시상의 외측배쪽핵 내측부(VLm), 배쪽전핵 거대세포부(VAmc), 등쪽내측핵의 소세포부(MDpc)와 부수질판부(MDpl)로 투사되며, 창백시상섬유나 소뇌시상섬유와 중복되지 않게 종지하는 것이 특징이다
②흑색질피개섬유(nigrotegmental fiber):흑색질 그물부(SNr)에서 뇌간으로 이어진다. 흑색질에서 상구로 이어지는 경로는 조류나 파충류에서는 기저핵의 주출력경로이다
교뇌와 중뇌의 경계부에서 시작되어 적색핵의 아래쪽 끝부분까지 뻗어 있는 대뇌각교뇌핵(PPN)은 아세틸콜린(acetylcholine)을 신경전달물질로 함유한 콜린성신경원(cholinergic neuron, Ch5 세포군)이며 제뇌동물(decerebrate animal)에서 이 핵의 신경원을 자극하면 사지가 교대로 움직이는 반사적인 보행운동이 일어나기 때문에 중뇌보행영역(mesencephalic locomotor region)이라고도 한다.
흑색질 그물부의 원심성 신경원도 신경전달물질로 GABA를 함유하고 있다.
3.흑색질 치밀부 (SNc)
치밀부(SNc)에는 멜라닌색소(melanin pigment)가 세포질 내에 있는 큰 신경원들이 밀집되어 있으며, 이 세포들은 신경전달물질로 도파민(dopamine)을 함유하고 있다. 치밀부(SNc)의 세포들은 큰 다극세포(multipolar neuron)로 핵소체는 크고 뚜렷하며, 니슬소체도 크고 짙게 염색되며 뚜렷하다. 두세 개의 긴 수상돌기(dendrite)가 나타나기도 한다. 멜라닌색소는
생후 4년 이후에나 나타나기 시작하며 나이가 많아질수록 점차 증가한다. 이 부분은 설치류의 A9 세포군에 해당된다.
4.흑색질 치밀부(SNc)의 연결
㈀구심성 연결 (afferent connection)
흑색질 치밀부(SNc)에는 창백핵의 내핵(GPi), 외핵(GPe) 두 부분 모두에서 기원되는 창백흑색질섬유(pallidonigral fiber)가 종지한다. 선조에서도 직접 연결되는 신경섬유가 있다. 선조에서 흑색질 치밀부로 직접 연결되는 선조신경원은 선조소체(striosome)에 있으며, 일부 변연피질(limbic cortex)에서 구심섬유를 받는다고 알려져 있다. 흑색질 치밀부는 대뇌피질에서 구심섬유를 받는다는 보고도 있다
㈁원심선 연결 (efferent connection)
흑색질 치밀부(SNc)의 도파민함유신경원은 주로 선조(striatum)로 원심섬유(흑색선조섬유 nigrostriatal fiber)를 보낸다. 이 섬유다발은 부위별 대응연결을 하고 있다. 흑색선조섬유는 선조의 제2형무극세포(A II)를 억제하는 작용을 한다고 알려져 있다. 흑색선조섬유가 기원되는 모든 신경원이 도파민을 함유하고 있지는 않으며, 약 20% 정도는 도파민을 함유하고 있지 않다고 알려져 있다. 선조(striatum)로 투사되는 흑색질 치밀부의 도파민성원심섬유를 선조관련도파민계(mesostriatal dopaminergic system)라고도 한다.
시상밑핵 (Subthalamic Nucleus)
시상밑핵(subtahalamic nucleus, STN)은 양쪽이 모두 볼록한 볼록렌즈 형태의 핵으로 간뇌(diencephalon)에서 파생된 구조이다. 배쪽 외측으로는 내섬유막(internal capsule)이 있고, 배쪽 내측으로는 렌즈핵다발(lenticular fasciculus)의 신경섬유가 지나간다. 위쪽으로는 포렐 H2 구역(Forel's field H2)을 사이에 두고 불확정구역(zona incerta)과 닿아있다. 흑색질의 위쪽 끝부분의 등쪽 외측에 있으며, 적색핵의 외측에 위치한다.
1.시상밑핵(STN)의 연결
㈀구심성 연결 (afferent connection)
시상밑핵(STN)은 주로 창백외핵(GPe)에서 구심섬유(창백시상밑핵섬유 pallidosubthalamic fiber)를 받는다. 입력부인 선조로 투사섬유를 보내지 않고 출력부인 창백으로 투사섬유를 보내는 구조는 시상밑핵이 유일하다
일차운동영역(M I), 전운동영역(premotor area) 및 전전두엽피질(prefrontal cortex)을 포함한 대뇌피질, 시상의 중심정중핵-다발옆핵 복합체(CM- PF complex), 중뇌의 대뇌각교뇌핵(PPN)에서도 구심섬유가 들어온다고 보고되어 있다
㈁원심성연결
배쪽선조(Ventral Striatum)와 배쪽창백(Ventral Pallidum)
선조체(corpus striatum)의 배쪽에는 경계가 명확하지 않은 핵군이 있으며 이 부위를 전뇌기저부(basal forebrain)라고 한다. 이 부분은 전관통질(anterior perforating substance)의 심부에 있는 회색질인 무명질(substantia innominata)과 그 앞쪽에 위치하는 중격의지핵(nucleus accumbens, nucleus accumbens septi) 및 그 주위부분을 말한다. 전관통질의 앞쪽 경계에는 후각삼각과 내측, 외측후각선조가 있고, 외측으로는 전이상엽피질(prepyriform cortex)과 편도체주위피질(periamygdaloid cortex)이 있다. 뒤쪽 경계는 브로카대각선조(diagonal band of Broca)와 시각로(optic tract), 시각신경교차(optic chiasm)로 이루어져 있다. 브로카대각선조는 중격부에서 기원되는 섬유다발로, 전관통질의 뒤쪽에까지 뻗어 있는 신경로이다. 전관통질의 표면은 내경동맥(internal carotid artery)이 전대뇌동맥(anterior cerebral artery)과 중간대뇌동맥(middle cerebral artery)으로 나누어지는 부분이며, 여기에서 나오는 수많은 중심가지(central branch)들이 이 부분을 뚫고 대뇌로 들어가기 때문에 관통질이라고 불리운다. 후각삼각의 바로 뒷부분 즉, 전관통질의 앞부분은 후각이 발달된 동물에서는 뚜렷하게 융기되어 있어 후각결절(olfactory tubercle)이라고 한다. 사람에서 후각결절 부위는 뚜렷하지 않다.
최근에는 대뇌기저부(basal forebrain area)에 있는 중격의지핵(nucleus accumbens)과 무명질(substantia innominata)의 기저핵에 포함시킨다. 이들을 이루는 세포의 형태와 연결이 선조체와 매우 비슷하며, 특히 중격의지핵은 선조의 배쪽 연장이라고 생각되므로 배쪽선조(ventral striatum)라 하고, 무명질의 일부는 창백핵의 배쪽 연장이라 생각되므로 배쪽창백(ventral pallidum)이라고 한다. 반면 선조와 창백은 등쪽선조(dorsal striatum)와 등쪽창백(dorsal pallidum)이라고 한다.
중격의지핵(nucleus accumbens, nucleus accumbens septi)은 미상핵 머리(head of caudate nucleus)와 조가비핵(putamen)의 배쪽에 있으며 두 핵이 내섬유막 아래에서 이어지는 부분이다. 이 핵 신경세포의 형태는 선조와 매우 비슷하여 중간크기의 세포들이 밀집되어 있는 형태로 나타난다.
중격의지핵의 배쪽 뒤쪽에는 무명질이 있다. 무명질에는 마이네르트기저핵(basal nucleus of Meynert)이 있으며 이 부분의 신경원은 아세틸콜린(acetylcholine)을 함유하고 있고 알츠하이머병(Altzheimer's disease)에서 현저한 변화가 나타나는 부위이다. 마이네르트기저핵의 신경원은 매우 크며, 아세틸콜린(acetylcholine)을 신경전달물질로 함유하고 있다. 이 핵은 메술램(Mesulam)의 Ch4 세포군에 해당되는 부위로 아세틸콜린성 신경핵 중 가장 크고 뚜렷하다. 중격핵에 있는 Ch1-Ch3 세포군과 함께 거대세포기저내측종뇌핵군(magnocellular basomedial telencephalic nuclear group)이라는 명칭으로 불리운다.
무명질에서 마이네르트기저핵을 제외한 비콜린성 부위는 창백핵과 이어져 있고, 신경세포의 형태와 연결도 창백핵과 비슷하기 때문에 이를 배쪽창백(ventral pallidum)이라고 한다.
배쪽창백과 배쪽선조는 전뇌기저부에 속하는 구조로 변연계(limbic system)로 분류하는 경우도 있지만, 대부분의 경우에는 기저핵(basal ganglia)의 일부로 분류한다. 이 부분은 기저핵의 일부로 변연계와 고도로 연결되어 있기 때문에 변연기저핵(limbic basal ganglia)이라고도 불리운다.
1.배쪽선조(ventral striatum)와 배쪽창백(ventral pallidum)의 연결
기저핵에서 이 부분은 주로 변연계(limbic system)와 연결되어 있으며, 체운동기능(somatic motor function)에 관계된 부분에 비해 복잡하게 연결되어 있다. 선조-창백핵에서 같이 배쪽선조(ventral striatum)가 입력부이며 배쪽창백(ventral pallidum)이 출력부이다
㈀배쪽선조(ventral striatum)
역시 선조와 마찬가지로 대뇌피질, 시상, 편도체(amygdaloid body), 뇌간의 도파민함유신경원, 솔기핵 등에서 구심섬유를 받는다. 다만 대뇌피질은 주로 변연피질(limbic cortex)이며, 도파민함유신경섬유는 선조와는 달리 배쪽피개구역(ventral tegmental area, A8)에서 주로 기원한다.
배쪽선조, 즉 중격의지핵으로 투사되는 대뇌피질부위는 내측전뇌피질(medial frontal cortex), 내후각뇌피질(entorhinal cortex), 해마형성체(hippocampal formation)의 해마이행부(subiculum)이며, 해마이행부에서의 연결은 부위별 대응연결을 하고 있어 해마이행부의 앞쪽 부분은 중격의지핵의 내측으로 투사되고 뒤쪽부분은 중격의지핵의 외측부분으로 투사된다. 변연피질에서 중격의지핵으로 연결되는 피질선조섬유(cortico-striate fiber)의 신경전달물질은 흥분성인 글루탐산(glutamic acid)이나 아스파라긴산(aspartic acid)이다. 시상에서는 중간핵군(midline nuclear group)에 속하는 맥락끈옆핵(paratenial nucleus)에서 구심성섬유를 받는다.
배쪽선조로 들어오는 편도체선조섬유(amygdalostriate fiber)는 주로 기저편도체핵(basal amygdaloid nucleus)에서 기원되며 피질선조섬유와 같이 부위별 대응연결을 하고 있다.
배쪽피개구역(ventral tegmental area, A8)에서 기원되는 도파민함유신경섬유는 선조에도 일부 종지하지만 대부분은 중격의지핵으로 이어진다. 반면 흑색질치밀부(SNr)에서는 거의 구심섬유가 들어오지 않는다.
배쪽선조의 원심섬유는 상당 부분이 배쪽창백으로 이어지지만, 주로 기저핵의 출력부로만 이어지는 선조의 원심섬유와는 달리 변연계의 다른 부분(중격핵, 시상하부, 배쪽피개구역, 솔기핵, 중뇌수도관주위회색질 등)으로 이어지는 원심섬유도 많다. 배쪽선조에서 배쪽창백으로 이어지는 선조창백섬유도 신경전달물질로 감마아미노부티르산(GABA)과 섭스탄스 피(substance P), 엔케팔린(enkephalin) 및 다이놀핀(dynorphin)을 함유하고 있다. 이들 중 엔케팔린(enkephalin)성 섬유는 등쪽창백으로도 이어지지만 섭스탄스 피(substance P)를 함유하는 신경종말은 배쪽창백에만 종지한다.
㈁배쪽창백(ventral pallidum)
주로 배쪽선조에서 구심성섬유를 받지만, 배쪽선조로 연결되는 거의 모든 부위(변연피질, 시상 중간핵군, 편도체, 배쪽피개구역 등)에서 구심섬유를 받는다.
원심섬유는 시상의 등쪽내측핵(MD)과 중간핵군(midline nuclei)을 경유하여 전전두엽피질(prefrontal cortex)로 연결되는 경로가 주경로이지만, 흑색질치밀부(SNr)와 배쪽피개구역(VTA) 및 대뇌각교뇌핵(pedunculopontine nucleus)으로도 이어진다.
기저핵의 연결회로
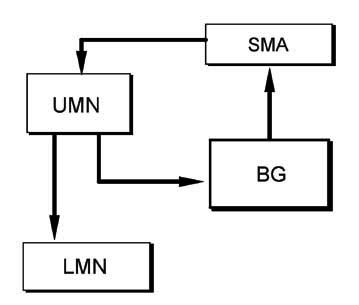
기저핵의 원심성연결은 주로 시상(thalamus)의 VL-VA 핵복합체-보완운동영역(supplementary motor area, SMA)-일차운동피질(M I)로 이어지며, 피라미드로(pyramidal tract)를 통해 하위운동신경원(lower motor neuron)에 영향을 준다. 또한 일차운동영역에서는 기저핵으로도 원심섬유를 보낸다. 따라서 기저핵의 회로는 운동피질에서 나가는 피라미드로(pyramidal tract)의 한 부회로(subcircuit)라고 말할 수 있으며 기저핵의 손상으로 나타나는 여러 가지 불수의적 운동은 피라미드로를 절단하면 모두 없어지게 된다.

BG 기저핵(basal ganglia)
SMA 보완운동영역(supplementary motor area)
UMN 상위운동신경원(upper motor neuron)
LMN 하위운동신경원(lower motor neuron)

기저핵의 연결을 자세히 살펴보면 대부분이 어느 한 부분에서 시작되어 다른 부분을 거쳐 다시 시작된 곳으로 되돌아오는 회로(circuit)을 구성하고 있다는 것을 알 수 있다. 이러한 회로들은 여러 종류의 평행한 회로(parallel circuit)로 구성되어 서로 중복되지 않는 것이 특징적이다. 여기에 또한 여러가지 짧은 부회로(accessory circuit)가 있다.
기저핵의 주회로는 선조-출력부(창백 GP 과 흑색질 그물부 SNr)-시상(VL-VA)-운동피질(motor cortex)-선조를 잇는 회로(그림 9-13, 9-14)라고 생각되지만, 선조-창백(GP)-흑색질 치밀부(SNc)-선조를 연결하는 회로와 창백외핵(GPe)-시상밑핵(STN)-출력부(창백 GP 과 흑색질 그물부 SNr)를 잇는 회로 역시 주요한 부회로이다. 이 이외에 선조-창백(GP)-시상(CM-PF)-선조를 잇는 회로와 선조-창백(GP)-외측고삐핵(lateral habenular nucleus, LHN)-솔기핵(raphe)-선조를 잇는 회로, 출력부와 대뇌각교뇌핵(PPN)이 이루는 회로도 있다. 또한 이러한 회로 내의 신경원은 상당히 여러 가지의 신경전달물질을 함유하고 있다. 또한 선조에서 창백을 거치지 않고 직접 흑색질 치밀부(SNc)를 경유하여 선조로 돌아오는 회로도 있다.
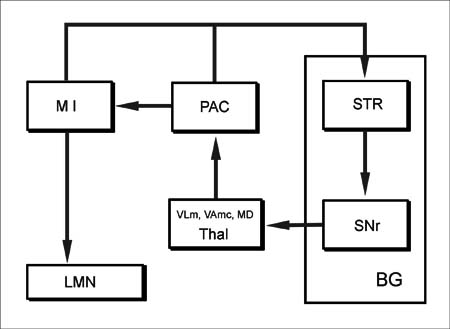
주회로인 선조-출력부-시상-운동피질을 잇는 회로는 다시 두 가지의 회로로 나눌 수 있다. 그 하나는 선조-창백내핵(GPi)-시상(VLo, VApc, CM)-보완운동영역(SMA)-일차운동영역(M I)-선조를 잇는 회로(그림 9-13) 는 선조-흑색질그물부(SNr)-시상(VLm, VAmc, MD)-전전두엽피질(prefrontal cortex)-일차운동영역(M I)을 잇는 회로(그림 9-14)이다. 창백(GP)에서 나가는 창백시상섬유는 시상의 VLo, VApc, CM 에 종지하며, 흑색질 그물부(SNr)에서 나가는 섬유는 시상의 VLm, VAmc, MD 에 종지된다. 또한 이 두 회로의 입력과 출력에 약간의 차이가 있다. 선조-창백(GP)으로 이어지는 회로는 주로 조가비핵을 통해 전운동영역이나 운동-감각영역에서 입력을 받으며, 출력은 보완운동영역(SMA) 쪽으로 나간다. 이를 주회로(main loop)이라고 한다(그림 9-13). 반면 선조-흑색질 그물부(SNr)로 이어지는 회로는 주로 전전두엽의 연합피질(prefrontal association cortex)에서 미상핵(caudate nu cleus)을 거쳐 입력을 받으며, 출력은 역시 전전두엽 연합피질 쪽으로 나가게 된다. 이 회로는 전전두엽 '연합' 회로(prefrontal 'association' loop)라고 한다

BG 기저핵(basal ganglia)
STR 선조(striatum)
SNr 흑색질 그물부(pars reticulata, substantia nigra)
Thal 시상(thalamus)
PAC 전전두엽 연합영역(prefrontal association area)
M I 일차운동 영역(primary motor area)
LMN 하위운동신경원(lower motor neuron)
흑색질 그물부(SNr)와 시상(VLm, VAmc, MD)을 경유하는 기저핵의 회로(전전두엽 '연합' 회로prefrontal 'association' loop).

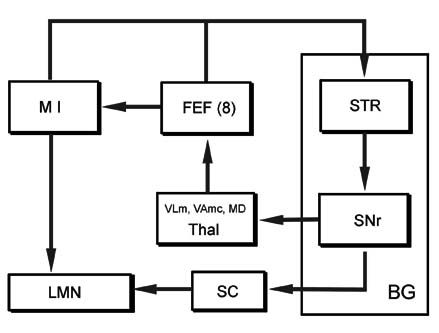
대뇌피질-기저핵-시상-대뇌피질을 잇는 회로는, 이 회로 이외에도 변연계의 일부인 대상이랑 앞부분(anterior cingulate gyrus)에서 주로 입력을 받아 배쪽선조-배쪽창백을 거쳐 시상의 등쪽내측핵(MD)으로 이어져 다시 변연계로 투사되는 회로도 있고, 전운동영역의 전안구운동영역(frontal eye field)에서 주로 입력을 받아 미상핵의 체부(body)-창백내핵(GPi)-흑색질 그물부(SNr)를 거쳐 다시 전안구영역으로 투사되는 안구운동회로(oculomotor loop)도 있다

BG 기저핵(basal ganglia)
STR 선조(striatum)
SNr 흑색질 그물부(pars reticulata, substantia nigra)
Thal 시상(thalamus)
FEF 전두엽안영역(frontal eye field)
M I 일차운동 영역(primary motor area)
SC 상구(superior colliculus)
LMN 하위운동신경원(lower motor neuron)
대뇌피질의 전안구영역(frontal eye field, 전두엽안구영역)-미상핵의 체부(body)-창백내핵(GPi)-흑색질 그물부(SNr)를 거쳐 다시 전안구영역으로 투사되는 안구운동회로(oculomotor loop). 일부 원심섬유는 상구(superior colliculus)로 연결되어 덮개척수로(tectospinal tract)를 통해 안구운동에 영향을 주기도 한다.

선조-흑색질 그물부(SNr)를 잇는 회로도 등쪽외측 전전두엽회로(dorsolateral prefrontal loop)와 외측 안와전두엽회로(lateral orbitofrontal loop)로 다시 나눌 수 있다
이와 같이 다양한 기저핵의 회로를 살펴보면 대뇌피질을 통해 기저핵으로 들어오는 입력이 기저핵회로를 경유하는 동안 여러 회로와 부회로를 거쳐 어떠한 선택적인 신경원의 단위집단(모듈 module)을 통해 출력이 이루어진다는 사실을 추측할 수 있다. 이러한 기저핵회로의 특성은 이미 프로그램화 되어 대뇌피질에 저장되어 있는 운동프로그램(motor program, motor routines)을 적절하게 선택(selection)하는데 있어 매우 유리하다고 생각된다. 즉 기저핵은 정상 상태에서 대뇌의 피질이나 시상 등 다른 여러 부위에서 오는 수많은 다양한 입력을 받아 이를 적절하게 처리한 후 그 상황에 가장 알맞는 운동프로그램을 선택하는 기능을 한다. 따라서 이 회로에 이상이 생기면 잘못된 운동프로그램의 선택으로 인하여 여러가지 불수의적 운동(involuntary movement)이 일어나거나 또는 운동프로그램을 적절하게 선택하지 못하여 운동불능(akinesia), 운동감소(hypokinesia), 운동완서(bradykinesia) 등의 증상이 일어날 수 있다고 생각된다. 또한 이러한 회로는 대뇌피질에 저장되어 있는 여러 운동프로그램의 단편(fragment)을 모아 하나의 운동프로그램으로 완성시키는데 있어서도 유리한 구조로 생각된다
기저핵의 손상에 따른 운동장애
기저핵의 일부 또는 전부가 손상되면 대체로 수의운동장애(dyskinesia)와 근경직(muscle rigidity)을 포함한 근긴장도와 자세의 이상(disturbance of posture and muscle tone)의 두 종류의 운동장애가 일어난다. 수의운동장애에는 자신의 의도와 관계없이 일어나는 불수의적 운동(involuntary movement)인 떨림(tremor, 진전), 아테토시스(무정위운동증, athetosis), 무도병(chorea), 발리즘(ballism) 등과 마비(paralysis)는 없는데도 불구하고 비정상적으로 운동의 시작이 안되거나 느려지는 운동감소증(hypokinesia), 운동불능증(akinesia), 운동완서(bradykinesia)등의 증상이 있으며, 근긴장도(muscle tone)의 이상은 근긴장항진(hypertonia) 즉 근경직(muscle rigidity)과 근긴장감소(hypotonia)로 나눌 수 있다.
1.떨림(tremor, 진전)
진폭(amplitude)과 주기(frequency)가 비교적 일정한 반복적인 운동이다. 기저핵 손상의 한 증상으로 나타나는 떨림은 수의운동을 하고 있지 않을 때 일어나며 수의운동이 일어날 경우에는 사라지는 휴지성 떨림(resting tremor)이다. 휴지성 떨림의 원인은 시상에서 기원되는 시상피질신경원(thalamocortical neuron)의 주기적인 활성 때문이라고 생각되고 있다. 이러한 경우 시상의 외측복측핵(VL)을 파괴하면 호전되는 경우가 많으며, 창백(GP)을 파괴하여도 이와 비슷한 결과가 오나 시상을 파괴하는 것 보다는 효과가 덜하다고 알려져 있다. 일차운동피질(M I)이나 추체로를 파괴하면 떨림은 없어지나 부전마비(paresis)가 온다. 파킨슨증후군의 주증상 중의 하나이며 간-렌즈핵변성(hepatolenticular degeneration, Wilson's disease)에서도 나타날 수 있다.
2.아테토시스(athetosis):
사지의 원위부(distal limb)를 불규칙하게 뒤틀거나 꿈틀거리는 느린 운동으로 대부분 근경직(muscle rigidity)이 동반되어 연속된 근경련(muscle spasm)의 형태로 나타난다. 대부분의 경우 선조(striatum)와 대뇌피질에 병변이 있으나 창백핵(GP)이나 시상(thalamus)의 병변으로 인해 유발되는 경우도 있다. 뇌성마비(cerebral palsy) 환자에서 흔히 볼 수 있다
3.무도병(chorea):
자신의 의지와는 관계없는 불수의적 운동이 마치 목적이 있는 운동처럼 활기 있고 우아한 연속적인 동작으로 나타난다. 대부분의 무도병에는 근긴장감소(hypotonia)가 동반되며, 주로 사지의 원위부와 얼굴의 표정근(muscles of expression!), 혀, 연하작용(deglutination)에 작용하는 근육 등에 잘 나타난다. 류마트열(rheumatic fever)의 합병증으로 올 수 있는 시드넘무도병(Sydenham's chorea)이나 유전적 질환인 헌팅튼병(Huntington's disease)의 대표적 증상의 하나로 주된 병변은 선조와 대뇌피질에 있다고 보고되어 있다
4.발리즘(ballism):격렬하게 사지를 휘두르는 불수의적 운동으로 상지대나 하지대 등 사지의 근위부 근육의 강력한 운동으로 나타난다. 이러한 경우 심한 근긴장감소(hypotonia)가 동반된다. 거의 대부분에서 시상밑핵(STN) 또는 시상밑핵과 연결된 주섬유다발이 손상된 경우에 나타나며, 흔히 손상 반대측에 일측성발리즘(hemiballism) 증상으로 나타난다.
5.운동불능증(akinesia)과 운동감소증(hypokinesia)
마비가 없음에도 불구하고 운동의 시작이 어렵거나 운동을 시작하는 빈도가 적어서 한 자세에서 다른 자세로의 이행이 곤란한 경우이며, 운동완서(bradykinesia)는 비정상적으로 운동이 느려지는 증상이다. 이들 모두는 파킨슨증후군에서 흔히 나타나는 증상이다.
6.근경직(muscle rigidity)
근육이 이완되지 않고 계속적으로 모든 근육이 수축하고 있는 상태로 일종의 근긴장항진증(hypertonia)이다. 근경직이 일어나는 원인에 대해서는 아직 잘 알려져 있지 않다. 근경직은 창백내핵(GPi)을 외과적으로 파괴하거나 시상의 VL 핵을 파괴하면 호전되며 척수의 후근을 잘라 척수로 들어가는 입력을 제거하거나 전각의 γ-운동신경원 활성을 억제하는 경우에도 감소된다고 알려져 있다. 근경직이 있는 경우에는 척수의 α-운동신경원과 γ-운동신경원에 대한 상척수성조절(supraspinal control)이 증가되고 강직성 신전반사(tonic stretch reflex)가 증가된다고 알려져 있으므로, 정상상태에서는 기저핵의 창백핵(GP)에서 나가는 억제성출력이 이를 감소시키지 않나 생각되고 있다. 근경직이 있는 관절을 억지로 펴려고 하면 마치 톱니바퀴가 돌아가는 것 같이 단계적으로 약간씩 펴지므로 이를 톱니바퀴현상(cogwheel phenomenon)이라고 한다.
반대로 근긴장감소(hypotonia)는 근육이 이완된 상태로 있는 것으로, 무도병과 발리즘에서는 근긴장감소가 동반된다.
7. 파킨슨증후군(Parkinson syndrome)
대표적인 증상으로 이 질환의 원인은 흑색질 치밀부(SNc) 신경원의 파괴와 이에 따른 도파민(dopamine)의 결핍에 있다
기저핵의 병변으로 인한 증상들은 같은 선조의 손상의 경우에도 어떤 경우에는 무도병이 나타나고 다른 경우에는 아테토시스 또 다른 경우에는 근긴장이상이 나타나는 등 거의 동일한 부위가 손상된 경우에도 다양하게 나타난다. 또한 선조, 창백(GP), 시상의 손상시 모두 근긴장이상이 나타나는 등 여러 다른 부위의 손상이 비슷한 증상으로 나타나기도 한다. 또한 동일한 질환 예를 들면 간-렌즈핵변성(Wilson's disease)의 경우, 파킨슨증후군 비슷한 운동불능-근경직증후군이 나타날 수도 있고, 아테토시스나 근긴장이상이 나타날 수도 있으며, 또는 이들이 혼합되어 나타나기도 한다. 일산화탄소 중독으로 인한 파킨슨증후군(Parkinson syndrome)이나 지발성운동장애(tardive dyskinesia)의 경우에는 증상의 발현이 그 원인에 비해 상당히 늦으며, 거의 동일한 부위의 손상에서도 어떤 경우에는 증상이 나타나고 어떤 경우에는 증상이 나타나지 않는 것으로 보아서는 손상이 있는 부위의 정확한 해부학적 범위 또는 손상된 신경원이 함유한 신경전달물질에 차이가 있거는 것으로 생각되며, 손상된 신경원의 재생(regeneration)과도 관계가 있는 것으로 추측된다.
기저핵의 연결회로와 기능이상으로 미루어 볼때 이 부분은 운동프로그램을 형성하고 선택(selection)하는 역할을 맡는 것으로 추측된다. 즉 기저핵은 정상 상태에서 대뇌의 피질이나 다른 여러 부위에서 오는 수많은 다양한 입력을 받아 이를 적절하게 처리한 후 그 상황에 가장 알맞는 운동프로그램을 선택하는 기능을 한다. 따라서 이 회로에 이상이 생기면 잘못된 운동프로그램의 선택으로 인하여 여러가지 불수의적 운동이 일어나거나 또는 운동프로그램을 적절하게 선택하지 못하여 운동불능, 운동감소, 운동완서 등의 증상이 일어나는것으로 생각된다.





















 Scientific American 'Intelligence'(Nov.1998) 표지에서
Scientific American 'Intelligence'(Nov.1998) 표지에서

































